大阪大学 大学院医学系研究科 Department of Anatomy and Neuroscience, Graduate School of Medicine, Osaka University
神経機能形態学講座(第二解剖)です。
研究内容Research Activity
ー脳はどう作られ、働くかー
1. 大脳における機能統合の基盤解明
脳機能、特に機能の統合の仕組みを解き明かすべく、長連合線維と呼ばれる、大脳皮質葉間(前頭葉と頭頂葉など)を結ぶ神経線維の回路形成の仕組みとその役割の解明に取り組んでいます。次の二つが主な取り組みです。
「大脳新皮質領野間回路の形成機構の解析」
「大脳新皮質ニューロンサブタイプに応じた回路形成機構の解析」
大脳新皮質には領野間回路だけでなく、領野内の局所回路や左右の大脳半球をつなぐ回路、視床や脊髄など大脳皮質以外の領域につながる回路などが存在しています。これらの回路を構成するニューロンは大脳新皮質で層を作って存在していますが、同じ層内に違うタイプのニューロンが同居していることもよくあります。さまざまなタイプのニューロンが、脳の発生の過程でどのようにそれぞれの個性を獲得し、それに基づいた固有の神経回路を形成するのか、そのメカニズムはまだあまりわかっていません。私たちは領野間回路と左右半球間回路のニューロンを比較しながら、個性獲得とタイプに応じた回路形成の機構を明らかにしようとしています。
2. 細胞移動による大脳皮質構築機構の解明
大脳(新)皮質の構成細胞は、グルタミン酸を神経伝達物質とする興奮性神経細胞が約8割を占め、残りはGABAを神経伝達物質とし、形態・機能的に多様な神経細胞で構成されます。これら神経細胞は、大脳皮質の外で生まれ、皮質内へと移入し、大脳皮質を構築します。特に、興奮性神経細胞は、大脳皮質深部にある側脳室の周囲にて生まれ、法線方向へ移動し、大脳皮質を形成します。この脳室周囲からの移動開始の調節機構は不明でしたが、我々は、脳室周囲に発現する新規分子FILIPを同定・クローニングし、FILIPがアクチン結合分子であるフィラミンの分解を促進することで細胞の移動性を低下させること、すなわち脳室周囲の細胞の移動開始は、負の制御系にてコントロールされることを世界に先駆け明らかにしました(Nagano et al., Nature Cell Biology,
2002. この内容は
Nature Reviews誌のハイライト欄にて紹介されました。Nature
Reviews: Molecular Cell Biology, 3:472-473, 2002)。さらに、神経細胞は移動途中で大きく形態を変え、その際、場合により移動方向を変えることが報告されていますが、この過程にフィラミンの発現量が大きく関わることを見い出しました(Nagano et al., J. Neurosci., 2004. この報告は同誌のハイライト論文に選出されました)。現在、FILIPのノックアウトマウスの解析を進めていますが、FILIPのノックアウトにより、大脳皮質内での細胞配置に異常があること、棘(スパイン)とよばれる神経伝達の場所であるシナプスにて働く構造物の形態が変化すること、フィラミン以外にも結合分子があり、その結合分子の働きを変化させていることなどを明らかとしました(現在投稿中)。また、移動途中で大きく細胞形態を変えると記しましたが、その過程が最終的な大脳皮質内での細胞配置に重要であること、細胞形態を変える過程に成長円錐におけるAbi2-WAVE2のリン酸化による制御が重要であることを見出しています(Xie et al., Cereb. Cortex, 2013)。
また、幾つかの大学と共同した研究も実施しています。札幌医科大を主とする共同研究として、大脳皮質脳室帯における細胞分化・脳室帯からの細胞移動にヒストンデアセチレースであるSIRT1が重要であることを示しました(Hisahara et al., Proc. Natl. Acad. Sci., USA,
2008)。私たちは、この研究では特に細胞分化と大脳皮質での細胞移動についての実験・解析を担当しましたが、この研究は掲載号の表紙に選出され、さらにCell誌においても注目すべき研究として紹介されました(2008年10月31号leading edge欄)。さらに、大阪市立大学を主とする共同研究において、カルパインが滑脳症の治療の鍵であることを示唆する成果を発表しています(Yamada et al., Nature Med, 2009)。
3. 大脳皮質から伸びる出力路の形成の仕組みの解明
大脳皮質から大脳皮質外(皮質下という)に伸びる神経線維の回路形成の分子レベルでの仕組みの解明に取り組んでいます。これは、障がいを受けた神経線維や、神経機能の回復にもつながることが期待されます。以下に詳細を記します。
大脳からの主要な出力路である皮質脊髄路、いわゆる錐体路は大脳皮質から脊髄へと投射し下位運動ニューロンを介して、体の運動機能をコントロールしています。錐体路起始細胞は大脳皮質第5層に存在し、皮質下へと長く伸びた軸索は最終的に脊髄まで達しますが、その主軸索が伸びた後に、いくつかの軸索からは上丘や橋核、下オリーブ核などの神経核へと枝分かれした軸索、いわゆる軸索側枝を伸ばすことが知られています。錐体路神経細胞は、脊髄運動ニューロンへと信号を送ると同時に、軸索側枝を介して複数の神経核へ信号を送ることで、より高度で精緻な運動機能を可能にしていると考えられます。特に、錐体路が進化的には新しい神経回路であり、ほ乳類で初めて出現し、ヒトで最も発達していることからも錐体路および軸索側枝の発達と高度な運動機能との関係がうかがえます。一方、成体脳においては、軸索側枝の形成は神経回路のつなぎ替えなどの可塑性に関与しているとされ、また神経が物理的・病的傷害を受けたときに、傷害領域へと軸索側枝を伸ばすことで代償機構としての役割を担うことが知られています。
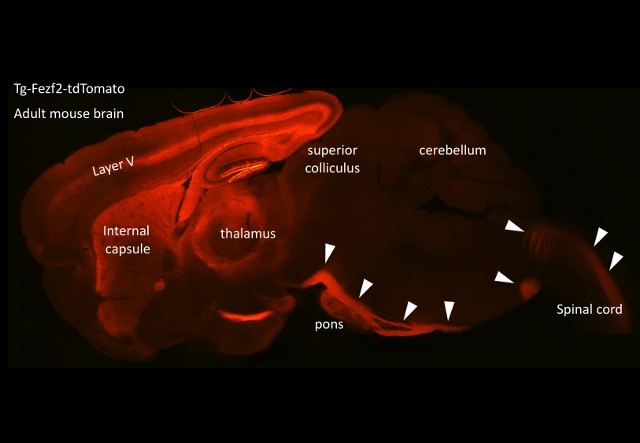
私たちは、マウス錐体路や任意の神経核神経細胞への遺伝子導入技術や、大脳皮質神経細胞の初代神経培養系、コラーゲンゲル内での組織培養法などの手法を用いて、この軸索側枝形成に関与する因子を探索すると共に、その因子に反応して側枝形成領域の軸索内部で起きるシグナル伝達及び細胞骨格の再編成といったイベントの解明に取り組んでいます。さらに、それらの知見に基づき、脳の可塑性における軸索側枝の形成機構とその役割の解明や、脳傷害に対する治療法への応用といった展開を目指しています。
4. 大脳皮質の発達の解明、発達の破たんによる疾病の本態の解明
発達障がいは、年々急増している病気ですが、責任遺伝子の多くはシナプスレベルでの情報伝達に関わるものの、患者さんの特徴的な病態との関係はいまだ謎で、そのため理解・治療が進んでいません。私たちは自閉症患者さんで大脳皮質領野間の機能分担に変化があることに着目し、シナプス機能に関わり、かつ自閉症にて大脳皮質領野間での発現が大きく変わる分子を手掛かりとして研究しています。すでに、そのモデルマウスを作製し、シナプスでの機能異常とマクロ的な回路機能変化との連関の解明を進めています。
5. シナプス部位での棘(スパイン)形成について
神経細胞が次の神経細胞に情報を伝達する場であるシナプスは、脳の情報伝達機構の基盤とされ、この部位での情報伝達効率の変化が、学習・記憶などの高次脳機能の細胞の実態と考えられています。シナプスは、一般に情報を伝える側(軸索末端)と情報を受け取る側(後シナプス構造)に分けられますが、特にスパイン(棘)に代表される後シナプス構造は、一般的に樹状突起フィロポディアがその形態を変化させ、形成されることが知られています。学習への関与の証拠として、実際に、神経細胞に一定の刺激を与えると、その細胞から次の神経細胞に情報が伝わるシナプス、特にスパインにおいて、動的にその形態・構造が大きく変化することが報告されています。この変化を長期増強(long-term potentiation; LTP)とよびますが、LTPの際には、さらに、スパインにおいて情報伝達に関わるグルタミン酸受容体の動態(数とスパイン膜表面に存在する分子数)が変化することが報告されています。
スパインのなかでも、シナプス後部の膜直下はシナプス後肥厚部(PSD)と呼ばれ、多種類のタンパク質が存在し、状況に応じて異なる複合体を形成し、情報伝達効率の調整に関わるとされています。中でも足場蛋白質PSD-95、カルシウム/カルモジュリン依存性リン酸化酵素II (CaMKII)は、グルタミン酸受容体AMPA-Rや NMDA-Rとともに、LTPの中心的分子であることが報告されています。近年、シナプス後部の膜上での AMPA-Rの局在量が増加することが、スパインの形態のダイナミックな変化とともに、LTPを担う直接の仕組みであると考えられています。しかしながら、スパインのダイナミックな形態変化、AMPA-Rの局在量変化の分子的な仕組み、さらには両者の相互関係はその有無も含め、十分には解明されていませんでした。最近、リン脂質の一種であるPIP3 が、スパインでのAMPA-R および PSD-95 の樹状突起シャフトからスパイン内部への局在変化に対し重要な役割を果たしていると報告されましたが (Sasaki J et al., Nat. Neurosci, 2010)、このことは、何らかの分子を介して、PIP3がAMPA-RとPSD-95の時空間的調節を行っていることを示唆しています。
私たちは、リン脂質の一種であるPIP3と特異的に結合する分子の解析を進める過程で、この分子がAMPA-Rに結合し、そのスパイン内の局在を変化させることを見出しました。先ほど報告された分子の実体であると想定しています。さらに、この分子が、スパインの成熟に関わることも見出しました。現在、その分子のノックアウトマウスを作製し、解析も進めています。